Table of Links
2. Methods
2.1. Modeling Panspermia and Terraformation
2.2. Identifying the Presence of Terraformed Planets and 2.3. Software and Availability
3. Results
3.1. Panspermia can increase the correlation between planets’ compositions and positions
3.2. Likely terraformed planets can be identified from clustering
5. Acknowledgements and References
APPENDIX
APPENDIX
A. APPENDIX
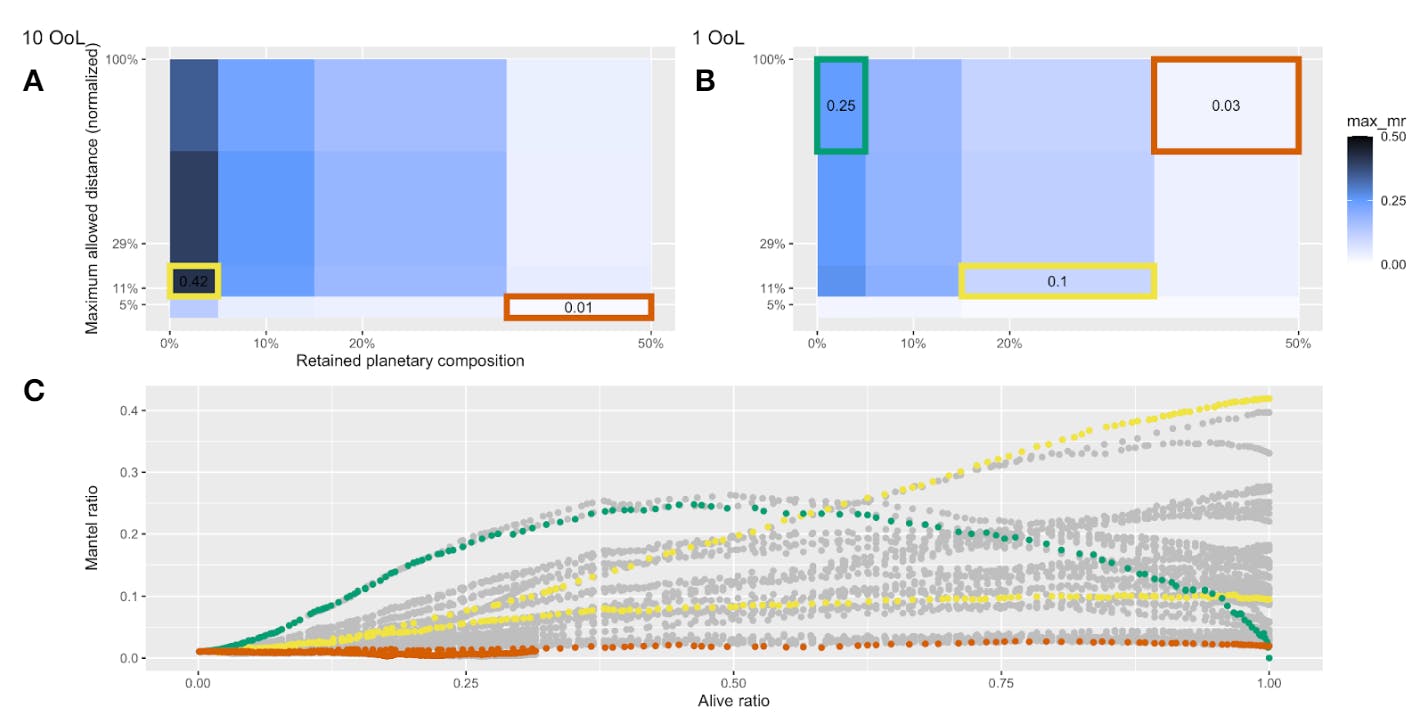
A.1. The mantel coefficient does not strictly increase with the proliferation of life
We investigated how changing the parameters of our model influenced the Mantel coefficient as a function of the ratio of terraformed planets. We find a few characteristic shapes of curves observed: flat, increasing then decreasing, and mostly increasing.
A.1.1. Additional Parameters
Mutation. Depending on the scenario, we include either a 0% or 10% chance of mutation for each element of the planet’s new composition, meaning that in the 10% scenario, with our length 10 composition vector, on average, 1 of the 10 elements is mutated during each terraformation event. Mutated elements are chosen from a continuous uniform random distribution of [0, 1], just like the initial compositions were chosen. Mutation is meant to represent the change in the kind of planetary observable characteristics which are compatible with life as life evolves over time.
Number of origins of life (OoL). Simulations are initialized with either 1 or 10 OoL (Fig. A2).
Compositions inherited from pre-terraformed planet. We vary the number of elements that the post-terraformed planet inherits from the pre-terraformed planet, n ∈ [0, 1, 2, 5], with the rest of the composition inherited from the incoming life.
A.1.2. Flat curve

A.1.3. Increasing then decreasing curve
When simulations are initialized with a single OoL, the progression of planet terraformation over time leads to an increasing then decreasing shaped Mantel coefficient curve, where the Mantel coefficient peaks after some of the planets have been terraformed, and then decreases afterwards (Fig. A1, data in green). Because the Mantel coefficient is measuring the correlation between positional and compositional distance matrices of planets, this correlation will decrease if either distance matrix becomes too homogeneous. With a single OoL, this is exactly what begins to happen with the compositional distance matrix, with the decline especially pronounced in the scenario with perfect replication (Fig. A1B, green box).
A.1.4. Mostly increasing curve













Authors:
(1) Harrison B. Smith, Earth-Life Science Institute, Tokyo Institute of Technology, Ookayama, Meguro-ku, Tokyo, Japan, and Blue Marble Space Institute of Science, Seattle, Washington, USA ([email protected]);
(2) Lana Sinapayen, Sony Computer Science Laboratories, Kyoto, Japan and National Institute for Basic Biology, Okazaki, Japan ([email protected]).
This paper is available on arxiv under CC BY-NC-ND 4.0 Deed license.